Are you looking for the best electronic dog collars in Australia?
Look no further! Here at Hidden Fence, we understand the importance of finding the right training tools for your furry friend. That’s why we’ve compiled a list of the top 3 electronic dog collars in Australia, each offering quality, value for money, intuitive, user-friendly design, and excellent training outcomes for dog owners. Let’s explore these top picks and discover how they can help in socialising your dog, teaching them acceptable behaviour, and fostering a harmonious relationship with your family.
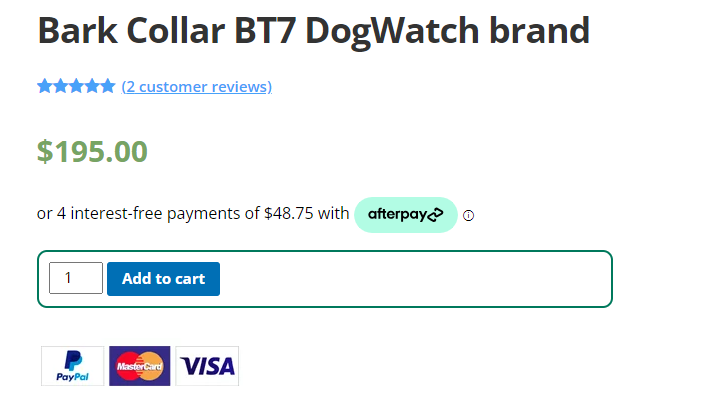
1. BT7 No Bark Training Collar
First on our list of top dog collars in Australia is the BT7 No Bark Training Collar. Perfect for teaching your dog to control their barking, this collar uses a combination of sound, vibration and static stimulus to gently remind your dog when it’s time to be quiet. The BT7 collar is specifically designed to be lightweight and comfortable, ensuring your dog won’t be hindered during their daily activities. This collar is an excellent tool for helping your dog understand when barking is inappropriate, leading to better behaviour and a more peaceful home environment.
LightweightWaterproofRechargeable15 Levels within 5 settings!Bark Counter TMVibration or e-StimulationProgressive or User set levelsLED Status3-Year warranty

No-Bark Trainer
The Bark Collar effectively trains your dog that there are limits to barking. Quickly & gently reduces and eliminates excessive barking behaviour.
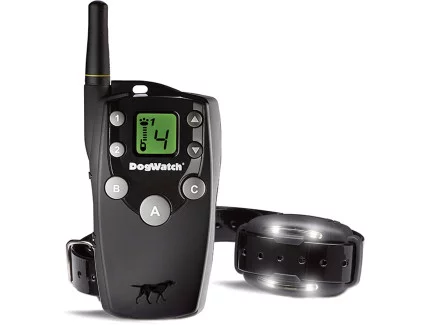
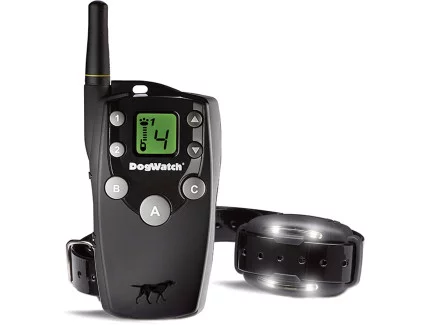
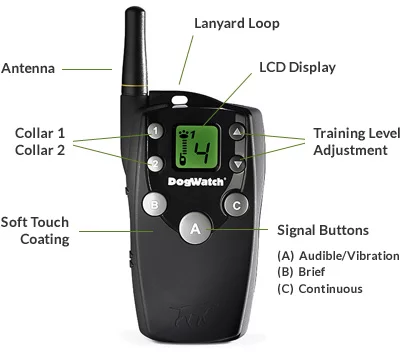
2. Big Leash Remote Control Training Collar
Next in our selection of top dog collars in Australia is the Big Leash Remote Control Training Collar. This innovative training tool offers unparalleled control and safety for your dog. With a range of features, including 15 levels of stimulation and a range of up to 800 meters, the Big Leash collar is ideal for teaching your dog various commands and obedience skills. Whether you’re in your backyard or out in the park, this collar ensures consistent training cues, helping your dog to learn faster and more effectively.
Features Include:
In-Touch™ Two-Way Communication15 Levels of Stimulation4 Signal OptionsFireFly® Nightlights800 Metre RangeTwo-Dog ReadyWaterproof ReceiverSoft-Touch™ Coating

Remote Trainer
Big Leash Remote Trainers allow you to communicate with your dog, so you can have more fun together. Helps you reinforce training commands and change unwanted behaviour.
3. SideWalker Trainer Collar
Last but certainly not least, in our roundup of the best dog collars in Australia is the SideWalker Trainer Collar. This collar is specifically designed to teach your dog proper leash manners. It delivers a gentle correction when your dog pulls on the leash, encouraging them to stay by your side. It’s an ideal solution for making walks more enjoyable for both you and your dog. The SideWalker collar is also adjustable, ensuring a comfortable fit for dogs of all sizes.
Digital Software5 adjustable settings, including vibrationProgressive levels within each settingWaterproof CollarLow battery indicator LEDLightweight1-Year Full Warranty

Walking Trainer
The SideWalker dog walking collar creates a more enjoyable walking experience for you and your dog. Gently corrects overly enthusiastic leash pulling and helps your dog learn appropriate on-leash behaviour.
Benefits of Electronic Dog Collars in Australia
Choosing the right dog collar in Australia is crucial for the effective training and socialisation of your dog. These top 3 electronic dog collars offer several benefits:
Enhanced Training: These collars provide consistent and intuitive training cues, making it easier for your dog to understand and follow commands.
Improved Socialisation: Properly trained dogs are more sociable and less likely to exhibit undesirable behaviours, leading to more positive interactions with other dogs and people.


Stronger Bond: Training your dog with these collars can lead to a stronger, more harmonious relationship between you and your pet.
Safety and Comfort: Designed with your dog’s comfort and safety in mind, these collars ensure that training is safe, consistent and effective.
When it comes to finding the best dog collars in Australia, the BT7 No Bark Training Collar, Big Leash Remote Control Training Collar, and SideWalker Trainer Collar stand out as the top choices. Not only do they offer exceptional quality and value, but they also provide dog owners with the tools necessary for successful training and a harmonious relationship with their pets. Explore these options and take the first step towards a well-behaved, happy dog today!